Никотинамидадениндинуклеотид
Никотинамидадениндинуклеотид | |
|---|---|
 | |
 | |
Общие | |
| Хим. формула | C₂₁H₂₇N₇O₁₄P₂ |
Физические свойства | |
| Состояние | белый порошок |
| Молярная масса | 663,43 г/моль |
Термические свойства | |
| Т. плав. | 160 ℃ |
Химические свойства | |
Растворимость в воде | 1 г/100 мл |
Классификация | |
| Номер CAS | 53-84-9 |
| PubChem | 5892 |
| ChemSpider | 5681 |
| Номер EINECS | 200-184-4 |
| RTECS | UU3450000 |
| ChEBI | 44215 |
| DrugBank | DB14128 |
SMILES | |
C1=CC(=C[N+](=C1)C2C(C(C(O2)COP(=O)([O-])OP(=O)(O)OCC3C(C(C(O3)N4C=NC5=C4N=CN=C5N)O)O)O)O)C(=O)N | |
InChI | |
1S/C21H27N7O14P2/c22-17-12-19(25-7-24-17)28(8-26-12)21-16(32)14(30)11(41-21)6-39-44(36,37)42-43(34,35)38-5-10-13(29)15(31)20(40-10)27-3-1-2-9(4-27)18(23)33/h1-4,7-8,10-11,13-16,20-21,29-32H,5-6H2,(H5-,22,23,24,25,33,34,35,36,37)/t10-,11-,13-,14-,15-,16-,20-,21-/m1/s1 | |
| Приводятся данные для стандартных условий (25 ℃, 100 кПа), если не указано иное. | |
Никотинамидадениндинуклеоти́д (англ. Nicotinamide adenine dinucleotide, сокр. NAD, НАД, устар. diphosphopyridine nucleotide, DPN, ДПН) — кофермент, имеющийся во всех живых клетках. NAD представляет собой динуклеотид и состоит из двух нуклеотидов, соединённых своими фосфатными группами. Один из нуклеотидов в качестве азотистого основания содержит аденин, другой — никотинамид. Никотинамидадениндинуклеотид существует в двух формах: окисленной (NAD+, NADox) и восстановленной (NADH, NADred).
В метаболизме NAD задействован в окислительно-восстановительных реакциях, перенося электроны из одной реакции в другую. Таким образом, в клетках NAD находится в двух функциональных состояниях: его окисленная форма, NAD+, является окислителем и забирает электроны от другой молекулы, восстанавливаясь в NADH, который далее служит восстановителем и отдаёт электроны. Такие реакции, сопряжённые с переносом электронов, являются основной сферой действия NAD. Однако NAD имеет и другие функции в клетке, в частности, он служит субстратом для ферментов, удаляющих или присоединяющих химические группы к белкам в ходе посттрансляционных модификаций. Из-за важности функций NAD, ферменты, участвующие в его метаболизме, являются мишенями для поиска новых препаратов[en].
В живых организмах NAD синтезируется de novo[en] из аминокислот аспартата или триптофана. Другие предшественники кофермента поступают в организм экзогенно, как, например, витамин ниацин (витамин В3) с пищей. Похожие соединения образуются в реакциях, приводящих к распаду NAD. После этого такие соединения проходят путь реутилизации, который возвращает их в активную форму. Некоторые молекулы NAD превращаются в никотинамидадениндинуклеотидфосфат (NADP). Этот близкий к NAD кофермент химически схож с ним, однако в метаболизме они выполняют разные функции.
Хотя NAD+ записывается с плюсом из-за формального положительного заряда атома азота, при физиологических значениях pH большая часть NAD+ на самом деле является анионом с отрицательным зарядом −1, а NADH — анионом с зарядом −2.
NAD называют «V-фактором», необходимым для роста гемофильной палочки (Haemophilus influenzae)[1].
Содержание
1 Физические и химические свойства
2 Концентрация и положение в клетках
3 Биосинтез
3.1 Образование de novo
3.2 Реутилизация
4 Функции
4.1 Оксидоредуктазы
4.2 Роль в окислительно-восстановительных реакциях
4.3 Другие внутриклеточные функции
4.4 Внеклеточные функции
5 Фармакологическое и медицинское применение
6 История
7 См. также
8 Примечания
9 Литература
Физические и химические свойства |
Никотинамидадениндинуклеотид состоит из двух нуклеотидов, соединённых мостиком из двух фосфатных групп, каждая из которых принадлежит одному из этих нуклеотидов. Кроме фосфатов, в состав этих нуклеотидов входит рибоза и азотистое основание, у одного нуклеотида оно представлено аденином, у другого — никотинамидом. Фосфаты прикрепляются к пятым атомам углерода (5′-положение), а азотистые основания — к первым (1′-положение). Никотинамид может присоединяться к аномерному 1′-атому в двух различных ориентациях, в связи с чем NAD существует в виде двух различных диастереомеров. В живых организмах встречается β-никотинамидный диастереомер NAD+[2].

Окисление и восстановление NAD
В метаболических процессах NAD участвует в окислительно-восстановительных реакциях, принимая или отдавая электроны[3]. Такие реакции, общее уравнение которых приводится ниже, включают формальную передачу гидрид-иона от исходного вещества (субстрата, RН2) к молекуле NAD+. При этом происходит нуклеофильное присоединение гидрида к никотинамидному фрагменту. Таким образом, исходное соединение RН2 окисляется до R, а NAD+ восстанавливается до NADH.
- RH2 + NAD+ → NADH + H+ + R.
Из электронной пары гидридного иона один электрон переносится на положительно заряженный азот в никотинамидном фрагменте, а атом водорода, оставшийся после отрыва электрона от гидридного иона, переносится на четвёртый атом углерода в кольце (С4), располагающийся напротив атома азота. Стандартный электродный потенциал окислительно-восстановительной пары NAD+/NADH составляет −0,32 вольт, что делает NADH сильным восстановителем[4]. Представленная выше реакция легко обратима, при этом NADH восстанавливает другую молекулу, а сам окисляется до NAD+. Поэтому кофермент может длительно циклично переходить из окисленного состояния в восстановленное, и наоборот, при этом расходования кофермента не происходит[2].
Физически обе формы кофермента представляют собой белый аморфный гигроскопичный порошок, хорошо растворимый в воде[5]. В твёрдом состоянии кофермент сохраняет стабильность в сухих условиях и в темноте. Раствор NAD+ бесцветен и сохраняет стабильность в течение недели при 4 °C и нейтральном pH, однако в щелочах и кислотах он быстро разрушается. При разложении NAD+ образуются продукты, являющиеся ингибиторами ферментов[6].
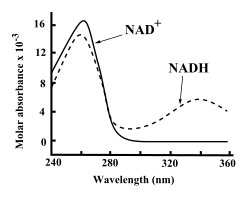
Спектры поглощения NAD+ и NADH
И NAD+, и NADH устойчиво поглощают ультрафиолетовое излучение из-за наличия аденина. Например, пик поглощения у NAD+ приходится на длину волны 259 нм, а коэффициент экстинкции составляет 16900 М−1см−1. NADH поглощает и волны больших длин, его второй пик поглощения ультрафиолета соответствует длине волны 339 нм, а коэффициент экстинкции равен 6200 М−1см−1[7]. Это различие в спектрах поглощения между окисленной и восстановленной формами кофермента позволяет простым образом измерить переход одной формы в другую при составлении характеристики активности фермента[en] путём измерения поглощения ультрафиолета при 340 нм с помощью спектрофотометра[7].
NAD+ и NADH флуоресцируют по-разному. В растворе NADH имеет пик эмиссии при 460 нм и продолжительность высвечивания 0,4 наносекунд, в то время как окисленная форма кофермента не флуоресцирует[8]. Параметры флуоресцирования NADH изменяются при связывании его с белками, поэтому эти изменения могут быть использованы для измерения константы диссоциации, которая широко используется при изучении кинетики ферментов[en][8][9]. Эти изменения во флуоресценции также могут применяться для оценки изменений в окислительно-восстановительном состоянии клетки методами флуоресцентной микроскопии[10].
Концентрация и положение в клетках |
В печени крысы суммарное количество NAD+ и NADH составляет приблизительно 1 мкмоль на грамм сырого веса, что в 10 раз больше концентрации NADP+ и NADPH в этих же клетках[11]. Реальную концентрацию NAD+ в цитозоле измерить сложнее, и, согласно современным представлениям, в клетках животных она составляет 0,3 мМ[12][13], а в клетках дрожжей приблизительно 1,0—2,0 мМ[14]. Однако более 80 % NADH, флуоресцирующего в митохондриях, находится в связанном виде, поэтому его концентрация в растворе значительно ниже[15].
Данные для других компартментов ограничены, хотя известно, что концентрация NAD+ в митохондриях схожа с таковой в цитозоле[13]. В митохондрию NAD+ из цитозоля проникает по специальным мембранным белкам-переносчикам[en], так как кофермент не может диффундировать сквозь мембраны[16].
Баланс между окисленной и восстановленной формой никотинамидадениндинуклеотида называется NAD+/NADH-отношением. Это отношение является важной частью т. н. окислительно-восстановительного состояния клетки — мерой и метаболической активности, и здоровья клетки[17]. Отношение NAD+/NADH имеет комплексное действие и оказывает влияние на активность ряда важнейших ферментов, среди которых глицеральдегид-3-фосфатдегидрогеназа[en] и пируватдегидрогеназный комплекс. В здоровых тканях млекопитающих отношение свободных NAD+ к NADH в цитоплазме обычно приблизительно равно 700; такое значение хорошо подходит для реакций окисления[18][19]. Общее отношение NAD+/NADH значительно ниже и составляет от 3 до 10 у млекопитающих[20]. В то же время отношение NADP+/NADPH в норме составляет около 0,005, то есть NADPH является преобладающей формой этого кофермента[21]. Различие в отношениях NAD+/NADH и NADP+/NADPH лежит в основе различных метаболических ролей NAD и NADP.
Биосинтез |
NAD+ синтезируется de novo из аминокислот, а также образуется путём реутилизации продуктов распада пиридиновых нуклеотидов.
Образование de novo |

Некоторые пути синтеза и потребления NAD+ у позвоночных. Пояснение сокращений см. в тексте
Большинство организмов синтезируют NAD+ из аминокислот[3]. Конкретный набор реакций отличается у различных организмов, однако для всех путей синтеза NAD+ характерно образование хинолината[en] (QA) из аспартата (многие бактерии и растения) либо триптофана (животные и некоторые бактерии)[22][23]. Хинолинат декарбоксилируется и фосфорибозилируется фосфорибозилпирофосфатом в никотинат-рибонуклеотид (NaMN). После этой стадии возможны альтернативные пути. В одном из таких путей происходит перенос аденилатного остатка с образованием адениндинуклеотида никотиновой кислоты (дезамино-NAD+, NaAD), после чего остаток никотиновой кислоты в составе NaAD амидируется с образованием никотинамидадениндинуклеотида[3].
На дополнительном этапе некоторые из новообразованных NAD+ превращаются в NADP+ферментом NAD+-киназой[en], которая фосфорилирует NAD+[24]. У большинства организмов этот фермент использует АТР в качестве донора фосфорильной группы, хотя некоторые бактерии, как, например, Mycobacterium tuberculosis и гипертермофильная архея Pyrococcus horikoshii[en] используют неорганический пирофосфат в качестве альтернативного донора фосфорильной группы[25][26].
Реутилизация |

Три основных предшественника NAD+
Кроме биосинтеза NAD+de novo из аминокислот аспартата или триптофана, клетки также способны образовывать NAD+ из готовой никотиновой кислоты и некоторых её производных. Хотя известны и другие предшественники, в этих метаболических путях обычно используются три природных соединения: никотиновая кислота (Na), никотинамид (Nam) и никотинамидрибозид (NR)[3]. Эти соединения могут попадать в организм экзогенно (например, с пищей, в которой содержится смесь никотиновой кислоты и никотинамида, называемая ниацином, или витамином В3). Однако эти соединения образуются и в самой клетке, где никотинамидный остаток высвобождается из NAD+ в реакциях переноса ADP-рибозных остатков. В самом деле, ферменты, обеспечивающие образование NAD+ из готовых производных никотиновой кислоты, сконцентрированы в ядре клетки, что может компенсировать большое количество реакций, протекающих в этой органелле с потреблением NAD+[27]. Клетки также могут получать NAD+ из своего внеклеточного окружения[28].
Несмотря на наличие пути синтеза NAD+de novo, реакции образования NAD+ из никотиновой кислоты и её производных жизненно важны для людей: при недостатке ниацина развивается заболевание пеллагра[29]. Такая высокая потребность в NAD+ обусловлена его постоянным расходованием в таких реакциях, как посттрансляционные модификации, поскольку переход NAD+ в NADH и обратно не изменяет общего количества кофермента[3].
Пути образования NAD+ из никотиновой кислоты и её производных у микроорганизмов отличаются от таковых у млекопитающих[30]. Некоторые патогены, например, дрожжи Candida glabrata[en] и бактерия Haemophilus influenzae ауксотрофны по NAD+ — они не способны синтезировать NAD+de novo, однако такие организмы, являясь зависимыми от экзогенных предшественников NAD+, могут синтезировать NAD+ путём реутилизации определённых производных никотиновой кислоты.[31][32]. У внутриклеточного патогена Chlamydia trachomatis отсутствуют какие-либо гены, которые потенциально могут быть вовлечены в пути образования и NAD+, и NADP+, и он должен получать оба этих кофермента извне[33].
Функции |

Россмановская укладка в лактатдегидрогеназе Cryptosporidium parvum[en]. NAD+ показан красным, бета-слои — жёлтым, альфа-спирали — пурпурным[34]
NAD выполняет несколько важнейших функций в метаболизме. Он выступает как кофермент в окислительно-восстановительных реакциях, как обязательный кофактор (простетическая группа) ферментов (циклаз фосфорилированных углеводов, различных эпимераз и др.), как донор ADP-рибозных остатков в реакциях ADP-рибозилирования[en] (одна из реакций посттрансляционной модификации белков), как предшественник циклической ADP-рибозы, являющейся вторичным посредником, а также как субстрат для бактериальных ДНК-лигаз и группы ферментов — сиртуинов, которые используют NAD+ для удаления ацетильных групп[en] с ферментов. Кроме этих метаболических функций, NAD+ может также выполнять важные функции вне клетки, так как он может выделяться из клетки спонтанно или в результате регулируемых процессов[35][36].
Оксидоредуктазы |
Наиболее важной функцией NAD+ в метаболизме является перенос электронов с одной молекулы на другую. Реакции такого типа катализируются большой группой ферментов, называемых оксидоредуктазами. Правильное название этих ферментов содержит название обоих их субстратов (окислителя и восстановителя), например, NADH-убихиноноксидоредуктаза катализирует перенос электронов с NADH на кофермент Q[37]. Однако, эти ферменты также называют дегидрогеназами и редуктазами: так, НАДН-убихиноноксидоредуктазу часто называют НАДН-дегидрогеназой или кофермент Q-редуктазой[38].
При связывании с белком NAD+ и NADH обычно располагаются в структурном мотиве[en] белка, известном как укладка Россмана[39]. Он был назван в честь Майкла Россмана[en], который был первым учёным, заметившим, что эта структура характерна для нуклеотид-связывающих белков[40]. В этом фолде имеются три или более параллельных бета-слоя, связанных двумя альфа-спиралями в порядке бета-альфа-бета-альфа-бета. В результате образуется общий бета-слой, с каждой стороны фланкированный слоем альфа-спиралей. Поскольку каждый фолд Россмана связывает лишь один нуклеотид, домены, связывающие динуклеотид NAD+, содержат два таких фолда, каждый из которых связывает один нуклеотид кофактора. Однако этот фолд не является универсальным среди NAD-зависимых ферментов; в частности, недавно был описан класс бактериальных ферментов, задействованных в метаболизме аминокислот, которые связывают NAD+, однако лишены этого мотива[41].
Связываясь с активным сайтом фермента, никотинамидный остаток NAD+ и субстрат взаимно ориентируются определённым образом, что благоприятствует эффективной передаче гидрида (H−). При изучении действия ферментов на дейтерированные субстраты было показано, что оксидоредуктазы селективно переносят гидрид к re- либо si-стороне никотинамидного остатка NAD+. В результате переноса на никотинамидный остаток D− вместо H− обазуется один из двух возможных диастереомеров NADH — это и позволяет установить, к какой именно стороне никотинамидного фрагмента NAD+ та или иная оксидоредуктаза переносит гидрид.

Высокая селективность обычно наблюдается также и в обратных процессах: оксидоредуктазы могут специфично переносить один из двух атомов водорода NADH (про-R либо про-S) к восстанавливаемому субстрату. Так, например, алкогольдегидрогеназа дрожжей и алкогольдегидрогеназы из печени человека, лошади переносят к субстрату про-R-атом водорода, а алкогольдегидрогеназа из Drosophila melanogaster производит восстановление при участии про-S-атома водорода[42]. Нативная алкогольдегидрогеназа дрожжей совершает одну «стереохимическую ошибку» на ~ 7 млрд актов катализа; показано, что мутации могут существенно снижать стереоспецифичность[43].

Эти факты нашли применение в исследованиях кинетики ферментативных реакций, а также в классификации ферментов. Оксидоредуктазы, взаимно ориентирующие субстраты таким образом, при котором гидрид атакует никотинамидный остаток с re-стороны (соответственно, в восстановленном коферменте подвижен HR), принято называть оксидоредуктазами класса A, тогда как в случае оксидоредуктаз класса B атака происходит с si-стороны (подвижен HS)[44].
При изучении ферментов, помимо описанной выше избирательности при выборе атома водорода в молекуле NADH, была обнаружена также и селективность по отношению к энантиотопным сторонам восстанавливаемого субстрата. Это указало на возможность использования ферментов в стереоселективном органическом синтезе для превращения кетонов в (R)- либо (S)-спирты.
Хотя механизмы связывания белков с NAD+ и NADP+ схожи, ферменты, как правило, демонстрируют высокую специфичность к NAD+ и NADP+[45]. Такая специфичность вытекает из различных метаболических ролей этих коферментов, и в их коферменто-связывающих сайтах располагаются различные наборы аминокислот. В частности, в активном центре NADP+-зависимых ферментов между аминокислотами основной цепочки и кислотно-фосфатной группой NADP+ образуется ионная связь, обусловленная определёнными зарядами аминокислотных остатков. В то же время в сайтах связывания с коферментом у NAD+-зависимых ферментов имеется другой набор зарядов аминокислот, что препятствует связыванию с NADP+. Впрочем, из этого общего правила существуют исключения: такие ферменты, как альдозоредуктаза[en], глюкозо-6-фосфатдегидрогеназа, метилентетрагидрофолатредуктаза у некоторых видов используют оба кофермента[46].
Роль в окислительно-восстановительных реакциях |
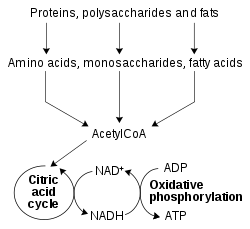
Опрощённая схема метаболизма с указанием ролей NAD+ и NADH
Окислительно-восстановительные реакции, катализируемые оксидоредуктазами, составляют важнейшую часть всех метаболических путей, однако наиболее значима их роль в процессах, связанных с выделением энергии из питательных веществ. В них такие восстановленные соединения, как глюкоза и жирные кислоты, окисляясь и в связи с этим выделяют энергию. Эта энергия запасается NAD+ при его восстановлении до NADH в ряде реакций β-окисления жирных кислот, гликолиза и цикла трикарбоновых кислот. У эукариот электроны, перенесённые на восстановленный в цитоплазме NADH, переносятся в митохондрию для восстановления митохондриальных NAD+ с помощью митохондриальных челночных механизмов[en], таких как малат-аспартатный челнок[en][47]. Митохондриальный NADH затем окисляется белками электроно-транспортной цепи, которые накачивают протоны в межмембранное пространство[en] из митохондриального матрикса, и благодаря энергии протонов в ходе окислительного фосфорилирования синтезируется ATP[48]. Такую же транспортную функцию челночные системы имеют и в хлоропластах[49].
Так как в этих связанных наборах реакций используются и окисленная, и восстановленная формы NAD, клетка поддерживает определённые концентрации NAD+ и NADH, и сохраняемое большое значение отношения NAD+/NADH позволяет этому коферменту выступать и в качестве окислителя, и в качестве восстановителя[50]. У NADPH, напротив, главной задачей является служить восстановителем в анаболических процессах, в частности, он вовлечён в такие процессы, как фотосинтез и синтез жирных кислот[en]. Поскольку NADPH выступает как сильный восстановитель и благодаря этому запускает окислительно-восстановительные реакции, значение отношения NADP+/NADPH поддерживается очень низким[50].
Несмотря на важную роль в катаболизме, NADH также участвует в некоторых анаболических процессах, например, глюконеогенезе[51]. Необходимость NADH в анаболических процессах создаёт проблему для микроорганизмов, растущих на питательных веществах, дающих лишь небольшое количество энергии. Например, нитрифицирующие бактерии Nitrobacter[en] окисляют нитрит до нитрата, и выделяющейся при окислении энергии достаточно для накачивания протонов и синтеза ATP, но не для непосредственного образования NADH[52]. Так как NADH всё-таки нужен в анаболических реакциях, эти бактерии используют фермент нитритоксидоредуктазу[en], которая создаёт достаточную протонодвижущую силу для того, чтобы заставить электроны двигаться по электроно-транспортной цепи в обратном направлении, что приводит к синтезу NADH[53].
Другие внутриклеточные функции |

ADP-рибоза
Кофермент NAD+ также расходуется в реакциях переноса ADP-рибозных[en] остатков. Например, ферменты ADP-рибозилтрансферазы[en] присоединяют свой ADP-рибозный остаток к белкам при посттрансляционной модификации, называемой ADP-рибозилированием[en][54]. ADP-рибозилирование может включать присоединение единственного ADP-рибозного остатка (моно(ADP-рибозил)ирование) или перенос ADP-рибозных остатков на белки с образованием длинных цепей из этих остатков (поли(ADP-рибозил)ирование)[55]. Первоначально моно-ADP-рибозилирование было известно как механизм созревания бактериальных токсинов, особенно холерного токсина, однако оно задействовано и в нормальной передаче сигналов между клетками[56][57]. Поли(ADP-рибозил)ирование осуществляется ферментами поли(ADP-рибозо)-полимеразами[55][58]. Поли(ADP-рибоз)ные цепи участвуют в регуляции некоторых клеточных процессов и особенно важны в клеточном ядре, где они задействованы в репарации ДНК и поддержании теломер[58]. Кроме внутриклеточных ADP-рибозилтрансфераз, недавно была описана группа внеклеточных ADP-рибозилтрансфераз, однако их функции пока неизвестны[59]. NAD+ также может присоединяться к клеточным РНК при 5′-терминальных модификациях[60].

Строение циклической ADP-рибозы
Другая функция NAD+ в передаче сигналов между клетками обусловлена тем, что он может служить предшественником для циклической ADP-рибозы — вторичного посредника, который образуется из NAD+ под действием ADP-рибозилциклаз[61]. Эта молекула участвует в кальциевых сигнальных путях[en], запуская высвобождение кальция из внутриклеточных депо[62]. Такое действие циклической ADP-рибозы обусловлено её связыванием и последующим открыванием кальциевых каналов, называемых рианодиновыми рецепторами[en]; эти рецепторы локализованы в мембранах органелл, например, эндоплазматического ретикулума[63].
NAD+ также используется при функционировании сиртуинов, например, Sir2[en][64]. Эти белки являются NAD-зависимыми деацетилазами. Их активность заключается в переносе ацетильных групп с субстратов-белков на ADP-рибозный остаток NAD+; это вызывает разрушение кофермента и высвобождение никотинамида и О-ацетил-ADP-рибозы. По-видимому, сиртуины участвуют в основном в регуляции транскрипции через деацетилирование гистонов и изменение структуры нуклеосом[65]. Однако сиртуины могут деацетилировать и негистоновые белки. Эта активность сиртуинов особенно интересна из-за их важной роли в регуляции старения[66].
Другими NAD-зависимыми ферментами являются бактериальные ДНК-лигазы, которые соединяют концы двух цепей ДНК, используя второй субстрат — NAD+ — как донор остатков AMP для присоединения к 5′-фосфату конца одной из цепей ДНК. Это промежуточное соединение далее атакуется 3′-гидроксильной группой конца другой цепи ДНК, и образуется новая фосфодиэфирная связь[67]. В отличие от бактериальных ДНК-лигаз, ДНК-лигазы эукариот используют ATP для образования промежуточных соединений ДНК-AMP[68].
Внеклеточные функции |
В последние годы было установлено значение NAD+ как внеклеточной сигнальной молекулы, участвующей в межклеточной коммуникации[36][69][70]. NAD+ выделяется нейросекреторными клетками[en][71] и из синаптосом[en] мозга[72] в кровеносные сосуды[35], мочевой пузырь[35][73], толстую кишку[74][75]. Предполагается, что NAD+ является новым нейромедиатором, который передаёт информацию от нейронов к эффекторным клеткам[en] в гладкомышечных органах[74][75]. Необходимы дальнейшие исследования для выяснения механизмов внеклеточных действий NAD+ и их влияния на здоровье и болезни человека.
Фармакологическое и медицинское применение |
Ферменты, вовлечённые в синтез и использование NAD+, имеют важное значение для фармакологии и исследований, направленных на поиск новых способов лечения болезней[76]. При разработке новых препаратов[en] NAD+ рассматривается с трёх позиций: как непосредственная мишень для лекарств, для разработки ингибиторов и активаторов ферментов, которые благодаря своей структуре изменяют активность NAD-зависимых ферментов и для изучения методов подавления биосинтеза NAD+[77].
В настоящий момент сам по себе кофермент NAD+ не используется для лечения каких бы то ни было заболеваний. Однако изучается его потенциальная роль в терапии нейродегенеративных заболеваний, таких как болезнь Альцгеймера и болезнь Паркинсона[3]. Имеются различные данные о действии NAD+ в нейродегенеративных болезнях. Некоторые исследования на мышах дают обнадёживающие результаты[78], однако клинические испытания на людях с использованием плацебо не дали какого-либо эффекта[79].
NAD+ также является непосредственной мишенью препарата изониазида, применяющегося для лечения туберкулёза — инфекции, вызываемой бактерией Mycobacterium tuberculosis. Изониазид является пролекарством и при попадании в клетку бактерии он активируется пероксидазой[en], которая окисляет это вещество в свободно-радикальную форму[80]. Этот радикал далее реагирует с NADH с образованием аддуктов, которые являются очень сильными ингибиторами ферментов редуктазы белка-переносчика еноил-ацила[en][81] и дигидрофолатредуктазы[82]. В одном эксперименте у мышей, которым давали NAD в течение недели, улучшалось взаимодействие клеточного ядра и митохондрий[83].
Из-за огромного количества оксидоредуктаз, использующих NAD+ и NADH в качестве субстратов и связывающихся с ними при помощи одного высококонсервативного структурного мотива, идея разработки ингибитора, блокирующего центр связывания NAD+, и специфичного лишь для определённого фермента, кажется сомнительной[84]. Однако это может быть выполнимым: так, ингибиторы, основанные на микофенолиновой кислоте[en] и тиазофурине[en], подавляют инозинмонофосфатдегидрогеназу[en] в сайте связывания с NAD+. Из-за важной роли этого фермента в метаболизме пуринов эти соединения могут быть полезными противораковыми и противовирусными препаратами или иммунодепрессантами[84][85]. Другие препараты являются не ингибиторами, а, наоборот, активаторами ферментов, вовлечённых в метаболизм NAD+. В частности, интересной мишенью для таких препаратов могут быть сиртуины, так как активация этих NAD-зависимых деацетилаз увеличивают продолжительность жизни[86]. Такие соединения, как ресвератрол, увеличивают активность этих ферментов, которые могут иметь большое значение благодаря их способности к переносу старения на более позднее время как у позвоночных[87], так и модельных организмов из числа беспозвоночных[88][89].
Из-за различий путей биосинтеза NAD+ у различных организмов, в частности, между бактериями и человеком, биосинтез NAD+ может стать новой сферой развития новых антибиотиков[90][91]. Например, фермент никотинамидаза[en], превращающая никотинамид в никотиновую кислоту, служит мишенью разрабатываемых лекарств, так как этот фермент отсутствует у человека, но имеется у бактерий и дрожжей[30].
История |

Артур Харден, один из первооткрывателей NAD+
Кофермент NAD+ был открыт английскими биохимиками Артуром Харденом и Уильямом Джоном Янгом[en] в 1906 году[92]. Они заметили, что добавление прокипячённого и профильтрованного экстракта дрожжей к непрокипячённым экстрактам значительно усиливало спиртовое брожение у последних. Неизвестный фактор, ответственный за это явление, они назвали коферментом. В ходе длительного и сложного выделения из экстрактов дрожжей этот теплостойкий фактор был идентифицирован как нуклеотид-сахарофосфат Хансом фон Эйлер-Хельпин[93]. В 1936 году немецкий учёный Отто Генрих Варбург установил функцию этого кофермента по переносу гидридного иона и определил, что в окислительно-восстановительных реакциях участвует никотинамидный остаток[94].
Источник никотинамида был определён в 1938 году, когда Конрад Элведжем[en] выделил ниацин из печени и показал, что этот витамин содержит никотиновую кислоту и никотинамид[95]. Позднее, в 1939 году, он предоставил первое убедительное доказательство того, что ниацин используется для образования NAD+[96]. В начале 1940-х Артур Корнберг сделал следующий шаг к пониманию роли NAD+ в метаболизме: он первым установил присутствие этого кофермента в биосинтетических путях[97]. Далее, в 1949 году американские биохимики Моррис Фридкин и Альберт Ленинджер доказали, что NAD+ связан с такими метаболическими путями, как цикл трикарбоновых кислот и окислительное фосфорилирование[98]. Наконец, в 1959 году Джек Присс (англ. Jack Preiss) и Филип Хандлер (англ. Philip Handler) описали ферменты и промежуточные соединения биосинтеза NAD+[99][100], поэтому путь синтеза NAD+de novo часто называют путём Присса — Хандлера в их честь.
Функции NAD и NADP, не связанные с окислительно-восстановительными реакциями, были открыты лишь в недавнее время[2]. Такой первой открытой функцией NAD+ было участие в качестве донора ADP-рибозного остатка в реакциях ADP-рибозилирования; это было установлено в начале 1960-х[101]. Более поздние исследования 1980-х и 1990-х годов показали участие NAD+ и NADP+ в передаче сигнала между клетками. В частности, действие циклической ADP-рибозы было установлено в 1987 году[102]. Метаболизм NAD+ и в XXI веке остаётся в сфере интенсивных исследований. Этот интерес особенно возрос после открытия в 2000 году Шинихиро Имаи (англ. Shinichiro Imai) и сотрудниками из Массачусетского технологического института NAD+-зависимых деацетилаз — сиртуинов[103].
См. также |
- NADP
- FAD
Примечания |
↑ X- AND V-FACTOR DISKS (неопр.).
↑ 123 Pollak N., Dölle C., Ziegler M. The power to reduce: pyridine nucleotides--small molecules with a multitude of functions. (англ.) // The Biochemical journal. — 2007. — Vol. 402, no. 2. — P. 205—218. — DOI:10.1042/BJ20061638. — PMID 17295611.
↑ 123456 Belenky P., Bogan K. L., Brenner C. NAD+ metabolism in health and disease. (англ.) // Trends in biochemical sciences. — 2007. — Vol. 32, no. 1. — P. 12—19. — DOI:10.1016/j.tibs.2006.11.006. — PMID 17161604.
↑ Unden G., Bongaerts J. Alternative respiratory pathways of Escherichia coli: energetics and transcriptional regulation in response to electron acceptors. (англ.) // Biochimica et biophysica acta. — 1997. — Vol. 1320, no. 3. — P. 217—234. — PMID 9230919.
↑ Windholz, Martha. The Merck Index: an encyclopedia of chemicals, drugs, and biologicals. — 10th. — Rahway NJ, US : Merck, 1983. — P. 909. — ISBN 0-911910-27-1.
↑ Biellmann J. F., Lapinte C., Haid E., Weimann G. Structure of lactate dehydrogenase inhibitor generated from coenzyme. (англ.) // Biochemistry. — 1979. — Vol. 18, no. 7. — P. 1212—1217. — PMID 218616.
↑ 12 Dawson, R. Ben. Data for biochemical research. — 3rd. — Oxford : Clarendon Press, 1985. — P. 122. — ISBN 0-19-855358-7.
↑ 12 Lakowicz J. R., Szmacinski H., Nowaczyk K., Johnson M. L. Fluorescence lifetime imaging of free and protein-bound NADH. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 1992. — Vol. 89, no. 4. — P. 1271—1275. — PMID 1741380.
↑ Jameson D. M., Thomas V., Zhou D. M. Time-resolved fluorescence studies on NADH bound to mitochondrial malate dehydrogenase. (англ.) // Biochimica et biophysica acta. — 1989. — Vol. 994, no. 2. — P. 187—190. — PMID 2910350.
↑ Kasimova M. R., Grigiene J., Krab K., Hagedorn P. H., Flyvbjerg H., Andersen P. E., Møller I. M. The free NADH concentration is kept constant in plant mitochondria under different metabolic conditions. (англ.) // The Plant cell. — 2006. — Vol. 18, no. 3. — P. 688—698. — DOI:10.1105/tpc.105.039354. — PMID 16461578.
↑ Reiss P. D., Zuurendonk P. F., Veech R. L. Measurement of tissue purine, pyrimidine, and other nucleotides by radial compression high-performance liquid chromatography. (англ.) // Analytical biochemistry. — 1984. — Vol. 140, no. 1. — P. 162—171. — PMID 6486402.
↑ Yamada K., Hara N., Shibata T., Osago H., Tsuchiya M. The simultaneous measurement of nicotinamide adenine dinucleotide and related compounds by liquid chromatography/electrospray ionization tandem mass spectrometry. (англ.) // Analytical biochemistry. — 2006. — Vol. 352, no. 2. — P. 282—285. — DOI:10.1016/j.ab.2006.02.017. — PMID 16574057.
↑ 12 Yang H., Yang T., Baur J. A., Perez E., Matsui T., Carmona J. J., Lamming D. W., Souza-Pinto N. C., Bohr V. A., Rosenzweig A., de Cabo R., Sauve A. A., Sinclair D. A. Nutrient-sensitive mitochondrial NAD+ levels dictate cell survival. (англ.) // Cell. — 2007. — Vol. 130, no. 6. — P. 1095—1107. — DOI:10.1016/j.cell.2007.07.035. — PMID 17889652.
↑ Belenky P., Racette F. G., Bogan K. L., McClure J. M., Smith J. S., Brenner C. Nicotinamide riboside promotes Sir2 silencing and extends lifespan via Nrk and Urh1/Pnp1/Meu1 pathways to NAD+. (англ.) // Cell. — 2007. — Vol. 129, no. 3. — P. 473—484. — DOI:10.1016/j.cell.2007.03.024. — PMID 17482543.
↑ Blinova K., Carroll S., Bose S., Smirnov A. V., Harvey J. J., Knutson J. R., Balaban R. S. Distribution of mitochondrial NADH fluorescence lifetimes: steady-state kinetics of matrix NADH interactions. (англ.) // Biochemistry. — 2005. — Vol. 44, no. 7. — P. 2585—2594. — DOI:10.1021/bi0485124. — PMID 15709771.
↑ Todisco S., Agrimi G., Castegna A., Palmieri F. Identification of the mitochondrial NAD+ transporter in Saccharomyces cerevisiae. (англ.) // The Journal of biological chemistry. — 2006. — Vol. 281, no. 3. — P. 1524—1531. — DOI:10.1074/jbc.M510425200. — PMID 16291748.
↑ Schafer F. Q., Buettner G. R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. (англ.) // Free radical biology & medicine. — 2001. — Vol. 30, no. 11. — P. 1191—1212. — PMID 11368918.
↑ Williamson D. H., Lund P., Krebs H. A. The redox state of free nicotinamide-adenine dinucleotide in the cytoplasm and mitochondria of rat liver. (англ.) // The Biochemical journal. — 1967. — Vol. 103, no. 2. — P. 514—527. — PMID 4291787.
↑ Zhang Q., Piston D. W., Goodman R. H. Regulation of corepressor function by nuclear NADH. (англ.) // Science (New York, N.Y.). — 2002. — Vol. 295, no. 5561. — P. 1895—1897. — DOI:10.1126/science.1069300. — PMID 11847309.
↑ Lin S. J., Guarente L. Nicotinamide adenine dinucleotide, a metabolic regulator of transcription, longevity and disease. (англ.) // Current opinion in cell biology. — 2003. — Vol. 15, no. 2. — P. 241—246. — PMID 12648681.
↑ Veech R. L., Eggleston L. V., Krebs H. A. The redox state of free nicotinamide-adenine dinucleotide phosphate in the cytoplasm of rat liver. (англ.) // The Biochemical journal. — 1969. — Vol. 115, no. 4. — P. 609—619. — PMID 4391039.
↑ Katoh A., Uenohara K., Akita M., Hashimoto T. Early steps in the biosynthesis of NAD in Arabidopsis start with aspartate and occur in the plastid. (англ.) // Plant physiology. — 2006. — Vol. 141, no. 3. — P. 851—857. — DOI:10.1104/pp.106.081091. — PMID 16698895.
↑ Foster J. W., Moat A. G. Nicotinamide adenine dinucleotide biosynthesis and pyridine nucleotide cycle metabolism in microbial systems. (англ.) // Microbiological reviews. — 1980. — Vol. 44, no. 1. — P. 83—105. — PMID 6997723.
↑ Magni G., Orsomando G., Raffaelli N. Structural and functional properties of NAD kinase, a key enzyme in NADP biosynthesis. (англ.) // Mini reviews in medicinal chemistry. — 2006. — Vol. 6, no. 7. — P. 739—746. — PMID 16842123.
↑ Sakuraba H., Kawakami R., Ohshima T. First archaeal inorganic polyphosphate/ATP-dependent NAD kinase, from hyperthermophilic archaeon Pyrococcus horikoshii: cloning, expression, and characterization. (англ.) // Applied and environmental microbiology. — 2005. — Vol. 71, no. 8. — P. 4352—4358. — DOI:10.1128/AEM.71.8.4352-4358.2005. — PMID 16085824.
↑ Raffaelli N., Finaurini L., Mazzola F., Pucci L., Sorci L., Amici A., Magni G. Characterization of Mycobacterium tuberculosis NAD kinase: functional analysis of the full-length enzyme by site-directed mutagenesis. (англ.) // Biochemistry. — 2004. — Vol. 43, no. 23. — P. 7610—7617. — DOI:10.1021/bi049650w. — PMID 15182203.
↑ Anderson R. M., Bitterman K. J., Wood J. G., Medvedik O., Cohen H., Lin S. S., Manchester J. K., Gordon J. I., Sinclair D. A. Manipulation of a nuclear NAD+ salvage pathway delays aging without altering steady-state NAD+ levels. (англ.) // The Journal of biological chemistry. — 2002. — Vol. 277, no. 21. — P. 18881—18890. — DOI:10.1074/jbc.M111773200. — PMID 11884393.
↑ Billington R. A., Travelli C., Ercolano E., Galli U., Roman C. B., Grolla A. A., Canonico P. L., Condorelli F., Genazzani A. A. Characterization of NAD uptake in mammalian cells. (англ.) // The Journal of biological chemistry. — 2008. — Vol. 283, no. 10. — P. 6367—6374. — DOI:10.1074/jbc.M706204200. — PMID 18180302.
↑ Henderson L. M. Niacin. (англ.) // Annual review of nutrition. — 1983. — Vol. 3. — P. 289—307. — DOI:10.1146/annurev.nu.03.070183.001445. — PMID 6357238.
↑ 12 Rongvaux A., Andris F., Van Gool F., Leo O. Reconstructing eukaryotic NAD metabolism. (англ.) // BioEssays : news and reviews in molecular, cellular and developmental biology. — 2003. — Vol. 25, no. 7. — P. 683—690. — DOI:10.1002/bies.10297. — PMID 12815723.
↑ Ma B., Pan S. J., Zupancic M. L., Cormack B. P. Assimilation of NAD(+) precursors in Candida glabrata. (англ.) // Molecular microbiology. — 2007. — Vol. 66, no. 1. — P. 14—25. — DOI:10.1111/j.1365-2958.2007.05886.x. — PMID 17725566.
↑ Reidl J., Schlör S., Kraiss A., Schmidt-Brauns J., Kemmer G., Soleva E. NADP and NAD utilization in Haemophilus influenzae. (англ.) // Molecular microbiology. — 2000. — Vol. 35, no. 6. — P. 1573—1581. — PMID 10760156.
↑ Gerdes S. Y., Scholle M. D., D'Souza M., Bernal A., Baev M. V., Farrell M., Kurnasov O. V., Daugherty M. D., Mseeh F., Polanuyer B. M., Campbell J. W., Anantha S., Shatalin K. Y., Chowdhury S. A., Fonstein M. Y., Osterman A. L. From genetic footprinting to antimicrobial drug targets: examples in cofactor biosynthetic pathways. (англ.) // Journal of bacteriology. — 2002. — Vol. 184, no. 16. — P. 4555—4572. — PMID 12142426.
↑ Senkovich O., Speed H., Grigorian A., Bradley K., Ramarao C. S., Lane B., Zhu G., Chattopadhyay D. Crystallization of three key glycolytic enzymes of the opportunistic pathogen Cryptosporidium parvum. (англ.) // Biochimica et biophysica acta. — 2005. — Vol. 1750, no. 2. — P. 166—172. — DOI:10.1016/j.bbapap.2005.04.009. — PMID 15953771.
↑ 123 Smyth L. M., Bobalova J., Mendoza M. G., Lew C., Mutafova-Yambolieva V. N. Release of beta-nicotinamide adenine dinucleotide upon stimulation of postganglionic nerve terminals in blood vessels and urinary bladder. (англ.) // The Journal of biological chemistry. — 2004. — Vol. 279, no. 47. — P. 48893—48903. — DOI:10.1074/jbc.M407266200. — PMID 15364945.
↑ 12 Billington R. A., Bruzzone S., De Flora A., Genazzani A. A., Koch-Nolte F., Ziegler M., Zocchi E. Emerging functions of extracellular pyridine nucleotides. (англ.) // Molecular medicine (Cambridge, Mass.). — 2006. — Vol. 12, no. 11-12. — P. 324—327. — DOI:10.2119/2006–00075.Billington. — PMID 17380199.
↑ Enzyme Nomenclature, Recommendations for enzyme names from the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (неопр.). Проверено 6 декабря 2007.
↑ NiceZyme View of ENZYME: EC 1.6.5.3 (неопр.). Expasy. Проверено 16 декабря 2007.
↑ Lesk A. M. NAD-binding domains of dehydrogenases. (англ.) // Current opinion in structural biology. — 1995. — Vol. 5, no. 6. — P. 775—783. — PMID 8749365.
↑ Rao S. T., Rossmann M. G. Comparison of super-secondary structures in proteins. (англ.) // Journal of molecular biology. — 1973. — Vol. 76, no. 2. — P. 241—256. — PMID 4737475.
↑ Goto M., Muramatsu H., Mihara H., Kurihara T., Esaki N., Omi R., Miyahara I., Hirotsu K. Crystal structures of Delta1-piperideine-2-carboxylate/Delta1-pyrroline-2-carboxylate reductase belonging to a new family of NAD(P)H-dependent oxidoreductases: conformational change, substrate recognition, and stereochemistry of the reaction. (англ.) // The Journal of biological chemistry. — 2005. — Vol. 280, no. 49. — P. 40875—40884. — DOI:10.1074/jbc.M507399200. — PMID 16192274.
↑ Chi-Huey Wong, G. M. Whitesides. Enzymes in Synthetic Organic Chemistry. — Oxford: Elsevier Science, 1994. — Т. 12. — С. 153—154. — 370 с. — (Tetrahedron Organic Chemistry). — ISBN 0080359426.
↑ Elmar G. Weinhold, Arthur Glasfeld, Andrew D. Ellington, and Steven A. Benner. Structural determinants of stereospecificity in yeast alcohol dehydrogenase (англ.) // Proceedings of the National Academy of Sciences U.S.A. : Научный журнал. — 1991. — Vol. 88, no. 19. — P. 8420—8424. — PMID 1924300.
↑ Bellamacina C. R. The nicotinamide dinucleotide binding motif: a comparison of nucleotide binding proteins. (англ.) // FASEB journal : official publication of the Federation of American Societies for Experimental Biology. — 1996. — Vol. 10, no. 11. — P. 1257—1269. — PMID 8836039.
↑ Carugo O., Argos P. NADP-dependent enzymes. I: Conserved stereochemistry of cofactor binding. (англ.) // Proteins. — 1997. — Vol. 28, no. 1. — P. 10—28. — PMID 9144787.
↑ Vickers T. J., Orsomando G., de la Garza R. D., Scott D. A., Kang S. O., Hanson A. D., Beverley S. M. Biochemical and genetic analysis of methylenetetrahydrofolate reductase in Leishmania metabolism and virulence. (англ.) // The Journal of biological chemistry. — 2006. — Vol. 281, no. 50. — P. 38150—38158. — DOI:10.1074/jbc.M608387200. — PMID 17032644.
↑ Bakker B. M., Overkamp K. M., van Maris A. J., Kötter P., Luttik M. A., van Dijken J. P., Pronk J. T. Stoichiometry and compartmentation of NADH metabolism in Saccharomyces cerevisiae. (англ.) // FEMS microbiology reviews. — 2001. — Vol. 25, no. 1. — P. 15—37. — PMID 11152939.
↑ Rich P. R. The molecular machinery of Keilin's respiratory chain. (англ.) // Biochemical Society transactions. — 2003. — Vol. 31, no. Pt 6. — P. 1095—1105. — DOI:10.1042/. — PMID 14641005.
↑ Heineke D., Riens B., Grosse H., Hoferichter P., Peter U., Flügge U. I., Heldt H. W. Redox Transfer across the Inner Chloroplast Envelope Membrane. (англ.) // Plant physiology. — 1991. — Vol. 95, no. 4. — P. 1131—1137. — PMID 16668101.
↑ 12 Nicholls DG. Bioenergetics 3. — 1st. — Academic Press, 2002. — ISBN 0-12-518121-3.
↑ Sistare F. D., Haynes R. C. Jr. The interaction between the cytosolic pyridine nucleotide redox potential and gluconeogenesis from lactate/pyruvate in isolated rat hepatocytes. Implications for investigations of hormone action. (англ.) // The Journal of biological chemistry. — 1985. — Vol. 260, no. 23. — P. 12748—12753. — PMID 4044607.
↑ Freitag A, Bock E (1990). “Energy conservation in Nitrobacter”. FEMS Microbiology Letters. 66 (1—3): 157—62. DOI:10.1111/j.1574-6968.1990.tb03989.x..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
↑ Starkenburg S. R., Chain P. S., Sayavedra-Soto L. A., Hauser L., Land M. L., Larimer F. W., Malfatti S. A., Klotz M. G., Bottomley P. J., Arp D. J., Hickey W. J. Genome sequence of the chemolithoautotrophic nitrite-oxidizing bacterium Nitrobacter winogradskyi Nb-255. (англ.) // Applied and environmental microbiology. — 2006. — Vol. 72, no. 3. — P. 2050—2063. — DOI:10.1128/AEM.72.3.2050-2063.2006. — PMID 16517654.
↑ Ziegler M. New functions of a long-known molecule. Emerging roles of NAD in cellular signaling. (англ.) // European journal of biochemistry / FEBS. — 2000. — Vol. 267, no. 6. — P. 1550—1564. — PMID 10712584.
↑ 12 Diefenbach J., Bürkle A. Introduction to poly(ADP-ribose) metabolism. (англ.) // Cellular and molecular life sciences : CMLS. — 2005. — Vol. 62, no. 7-8. — P. 721—730. — DOI:10.1007/s00018-004-4503-3. — PMID 15868397.
↑ Berger F., Ramírez-Hernández M. H., Ziegler M. The new life of a centenarian: signalling functions of NAD(P). (англ.) // Trends in biochemical sciences. — 2004. — Vol. 29, no. 3. — P. 111—118. — DOI:10.1016/j.tibs.2004.01.007. — PMID 15003268.
↑ Corda D., Di Girolamo M. Functional aspects of protein mono-ADP-ribosylation. (англ.) // The EMBO journal. — 2003. — Vol. 22, no. 9. — P. 1953—1958. — DOI:10.1093/emboj/cdg209. — PMID 12727863.
↑ 12 Bürkle A. Poly(ADP-ribose). The most elaborate metabolite of NAD+. (англ.) // The FEBS journal. — 2005. — Vol. 272, no. 18. — P. 4576—4589. — DOI:10.1111/j.1742-4658.2005.04864.x. — PMID 16156780.
↑ Seman M., Adriouch S., Haag F., Koch-Nolte F. Ecto-ADP-ribosyltransferases (ARTs): emerging actors in cell communication and signaling. (англ.) // Current medicinal chemistry. — 2004. — Vol. 11, no. 7. — P. 857—872. — PMID 15078170.
↑ Chen Y. G., Kowtoniuk W. E., Agarwal I., Shen Y., Liu D. R. LC/MS analysis of cellular RNA reveals NAD-linked RNA. (англ.) // Nature chemical biology. — 2009. — Vol. 5, no. 12. — P. 879—881. — DOI:10.1038/nchembio.235. — PMID 19820715.
↑ Guse A. H. Biochemistry, biology, and pharmacology of cyclic adenosine diphosphoribose (cADPR). (англ.) // Current medicinal chemistry. — 2004. — Vol. 11, no. 7. — P. 847—855. — PMID 15078169.
↑ Guse A. H. Regulation of calcium signaling by the second messenger cyclic adenosine diphosphoribose (cADPR). (англ.) // Current molecular medicine. — 2004. — Vol. 4, no. 3. — P. 239—248. — PMID 15101682.
↑ Guse A. H. Second messenger function and the structure-activity relationship of cyclic adenosine diphosphoribose (cADPR). (англ.) // The FEBS journal. — 2005. — Vol. 272, no. 18. — P. 4590—4597. — DOI:10.1111/j.1742-4658.2005.04863.x. — PMID 16156781.
↑ North B. J., Verdin E. Sirtuins: Sir2-related NAD-dependent protein deacetylases. (англ.) // Genome biology. — 2004. — Vol. 5, no. 5. — P. 224. — DOI:10.1186/gb-2004-5-5-224. — PMID 15128440.
↑ Blander G., Guarente L. The Sir2 family of protein deacetylases. (англ.) // Annual review of biochemistry. — 2004. — Vol. 73. — P. 417—435. — DOI:10.1146/annurev.biochem.73.011303.073651. — PMID 15189148.
↑ Trapp J., Jung M. The role of NAD+ dependent histone deacetylases (sirtuins) in ageing. (англ.) // Current drug targets. — 2006. — Vol. 7, no. 11. — P. 1553—1560. — PMID 17100594.
↑ Wilkinson A., Day J., Bowater R. Bacterial DNA ligases. (англ.) // Molecular microbiology. — 2001. — Vol. 40, no. 6. — P. 1241—1248. — PMID 11442824.
↑ Schär P., Herrmann G., Daly G., Lindahl T. A newly identified DNA ligase of Saccharomyces cerevisiae involved in RAD52-independent repair of DNA double-strand breaks. (англ.) // Genes & development. — 1997. — Vol. 11, no. 15. — P. 1912—1924. — PMID 9271115.
↑ Ziegler M., Niere M. NAD+ surfaces again. (англ.) // The Biochemical journal. — 2004. — Vol. 382, no. Pt 3. — P. e5–6. — DOI:10.1042/BJ20041217. — PMID 15352307.
↑ Koch-Nolte F., Fischer S., Haag F., Ziegler M. Compartmentation of NAD+-dependent signalling. (англ.) // FEBS letters. — 2011. — Vol. 585, no. 11. — P. 1651—1656. — DOI:10.1016/j.febslet.2011.03.045. — PMID 21443875.
↑ Yamboliev I. A., Smyth L. M., Durnin L., Dai Y., Mutafova-Yambolieva V. N. Storage and secretion of beta-NAD, ATP and dopamine in NGF-differentiated rat pheochromocytoma PC12 cells. (англ.) // The European journal of neuroscience. — 2009. — Vol. 30, no. 5. — P. 756—768. — DOI:10.1111/j.1460-9568.2009.06869.x. — PMID 19712094.
↑ Durnin L., Dai Y., Aiba I., Shuttleworth C. W., Yamboliev I. A., Mutafova-Yambolieva V. N. Release, neuronal effects and removal of extracellular β-nicotinamide adenine dinucleotide (β-NAD⁺) in the rat brain. (англ.) // The European journal of neuroscience. — 2012. — Vol. 35, no. 3. — P. 423—435. — DOI:10.1111/j.1460-9568.2011.07957.x. — PMID 22276961.
↑ Breen L. T., Smyth L. M., Yamboliev I. A., Mutafova-Yambolieva V. N. beta-NAD is a novel nucleotide released on stimulation of nerve terminals in human urinary bladder detrusor muscle. (англ.) // American journal of physiology. Renal physiology. — 2006. — Vol. 290, no. 2. — P. 486—495. — DOI:10.1152/ajprenal.00314.2005. — PMID 16189287.
↑ 12 Mutafova-Yambolieva V. N., Hwang S. J., Hao X., Chen H., Zhu M. X., Wood J. D., Ward S. M., Sanders K. M. Beta-nicotinamide adenine dinucleotide is an inhibitory neurotransmitter in visceral smooth muscle. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2007. — Vol. 104, no. 41. — P. 16359—16364. — DOI:10.1073/pnas.0705510104. — PMID 17913880.
↑ 12 Hwang S. J., Durnin L., Dwyer L., Rhee P. L., Ward S. M., Koh S. D., Sanders K. M., Mutafova-Yambolieva V. N. β-nicotinamide adenine dinucleotide is an enteric inhibitory neurotransmitter in human and nonhuman primate colons. (англ.) // Gastroenterology. — 2011. — Vol. 140, no. 2. — P. 608—617. — DOI:10.1053/j.gastro.2010.09.039. — PMID 20875415.
↑ Sauve A. A. NAD+ and vitamin B3: from metabolism to therapies. (англ.) // The Journal of pharmacology and experimental therapeutics. — 2008. — Vol. 324, no. 3. — P. 883—893. — DOI:10.1124/jpet.107.120758. — PMID 18165311.
↑ Khan J. A., Forouhar F., Tao X., Tong L. Nicotinamide adenine dinucleotide metabolism as an attractive target for drug discovery. (англ.) // Expert opinion on therapeutic targets. — 2007. — Vol. 11, no. 5. — P. 695—705. — DOI:10.1517/14728222.11.5.695. — PMID 17465726.
↑ Kaneko S., Wang J., Kaneko M., Yiu G., Hurrell J. M., Chitnis T., Khoury S. J., He Z. Protecting axonal degeneration by increasing nicotinamide adenine dinucleotide levels in experimental autoimmune encephalomyelitis models. (англ.) // The Journal of neuroscience : the official journal of the Society for Neuroscience. — 2006. — Vol. 26, no. 38. — P. 9794—9804. — DOI:10.1523/JNEUROSCI.2116-06.2006. — PMID 16988050.
↑ Swerdlow R. H. Is NADH effective in the treatment of Parkinson's disease? (англ.) // Drugs & aging. — 1998. — Vol. 13, no. 4. — P. 263—268. — PMID 9805207.
↑ Timmins G. S., Deretic V. Mechanisms of action of isoniazid. (англ.) // Molecular microbiology. — 2006. — Vol. 62, no. 5. — P. 1220—1227. — DOI:10.1111/j.1365-2958.2006.05467.x. — PMID 17074073.
↑ Rawat R., Whitty A., Tonge P. J. The isoniazid-NAD adduct is a slow, tight-binding inhibitor of InhA, the Mycobacterium tuberculosis enoyl reductase: adduct affinity and drug resistance. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2003. — Vol. 100, no. 24. — P. 13881—13886. — DOI:10.1073/pnas.2235848100. — PMID 14623976.
↑ Argyrou A., Vetting M. W., Aladegbami B., Blanchard J. S. Mycobacterium tuberculosis dihydrofolate reductase is a target for isoniazid. (англ.) // Nature structural & molecular biology. — 2006. — Vol. 13, no. 5. — P. 408—413. — DOI:10.1038/nsmb1089. — PMID 16648861.
↑ Gomes A. P., Price N. L., Ling A. J., Moslehi J. J., Montgomery M. K., Rajman L., White J. P., Teodoro J. S., Wrann C. D., Hubbard B. P., Mercken E. M., Palmeira C. M., de Cabo R., Rolo A. P., Turner N., Bell E. L., Sinclair D. A. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. (англ.) // Cell. — 2013. — Vol. 155, no. 7. — P. 1624—1638. — DOI:10.1016/j.cell.2013.11.037. — PMID 24360282.
↑ 12 Pankiewicz K. W., Patterson S. E., Black P. L., Jayaram H. N., Risal D., Goldstein B. M., Stuyver L. J., Schinazi R. F. Cofactor mimics as selective inhibitors of NAD-dependent inosine monophosphate dehydrogenase (IMPDH)--the major therapeutic target. (англ.) // Current medicinal chemistry. — 2004. — Vol. 11, no. 7. — P. 887—900. — PMID 15083807.
↑ Franchetti P., Grifantini M. Nucleoside and non-nucleoside IMP dehydrogenase inhibitors as antitumor and antiviral agents. (англ.) // Current medicinal chemistry. — 1999. — Vol. 6, no. 7. — P. 599—614. — PMID 10390603.
↑ Kim E. J., Um S. J. SIRT1: roles in aging and cancer. (англ.) // BMB reports. — 2008. — Vol. 41, no. 11. — P. 751—756. — PMID 19017485.
↑ Valenzano D. R., Terzibasi E., Genade T., Cattaneo A., Domenici L., Cellerino A. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short-lived vertebrate. (англ.) // Current biology : CB. — 2006. — Vol. 16, no. 3. — P. 296—300. — DOI:10.1016/j.cub.2005.12.038. — PMID 16461283.
↑ Howitz K. T., Bitterman K. J., Cohen H. Y., Lamming D. W., Lavu S., Wood J. G., Zipkin R. E., Chung P., Kisielewski A., Zhang L. L., Scherer B., Sinclair D. A. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. (англ.) // Nature. — 2003. — Vol. 425, no. 6954. — P. 191—196. — DOI:10.1038/nature01960. — PMID 12939617.
↑ Wood J. G., Rogina B., Lavu S., Howitz K., Helfand S. L., Tatar M., Sinclair D. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. (англ.) // Nature. — 2004. — Vol. 430, no. 7000. — P. 686—689. — DOI:10.1038/nature02789. — PMID 15254550.
↑ Rizzi M., Schindelin H. Structural biology of enzymes involved in NAD and molybdenum cofactor biosynthesis. (англ.) // Current opinion in structural biology. — 2002. — Vol. 12, no. 6. — P. 709—720. — PMID 12504674.
↑ Begley T. P., Kinsland C., Mehl R. A., Osterman A., Dorrestein P. The biosynthesis of nicotinamide adenine dinucleotides in bacteria. (англ.) // Vitamins and hormones. — 2001. — Vol. 61. — P. 103—119. — PMID 11153263.
↑ A. Harden, W. J. Young (24 October 1906). “The alcoholic ferment of yeast-juice Part II.--The coferment of yeast-juice”. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character. 78 (526): 369—375. JSTOR 80144.
↑ Fermentation of sugars and fermentative enzymes (неопр.) (PDF). Nobel Lecture, 23 May 1930. Nobel Foundation. Проверено 30 сентября 2007.
↑ Warburg O, Christian W. (1936). “Pyridin, der wasserstoffübertragende bestandteil von gärungsfermenten (pyridin-nucleotide)”. Biochemische Zeitschrift [нем.]. 287: 291. DOI:10.1002/hlca.193601901199. Неизвестный параметр|trans_title=(справка)
↑ Elvehjem CA, Madden RJ, Strong FM, Woolley DW (1938). “The isolation and identification of the anti-black tongue factor” (PDF). J. Biol. Chem. 123 (1): 137—49.
↑ Axelrod AE, Madden RJ, Elvehjem CA (1939). “The effect of a nicotinic acid deficiency upon the coenzyme I content of animal tissues” (PDF). J. Biol. Chem. 131 (1): 85—93.
↑ KORNBERG A. The participation of inorganic pyrophosphate in the reversible enzymatic synthesis of diphosphopyridine nucleotide. (англ.) // The Journal of biological chemistry. — 1948. — Vol. 176, no. 3. — P. 1475. — PMID 18098602.
↑ FRIEDKIN M., LEHNINGER A. L. Esterification of inorganic phosphate coupled to electron transport between dihydrodiphosphopyridine nucleotide and oxygen. (англ.) // The Journal of biological chemistry. — 1949. — Vol. 178, no. 2. — P. 611—644. — PMID 18116985.
↑ PREISS J., HANDLER P. Biosynthesis of diphosphopyridine nucleotide. I. Identification of intermediates. (англ.) // The Journal of biological chemistry. — 1958. — Vol. 233, no. 2. — P. 488—492. — PMID 13563526.
↑ PREISS J., HANDLER P. Biosynthesis of diphosphopyridine nucleotide. II. Enzymatic aspects. (англ.) // The Journal of biological chemistry. — 1958. — Vol. 233, no. 2. — P. 493—500. — PMID 13563527.
↑ CHAMBON P., WEILL J. D., MANDEL P. Nicotinamide mononucleotide activation of new DNA-dependent polyadenylic acid synthesizing nuclear enzyme. (англ.) // Biochemical and biophysical research communications. — 1963. — Vol. 11. — P. 39—43. — PMID 14019961.
↑ Clapper D. L., Walseth T. F., Dargie P. J., Lee H. C. Pyridine nucleotide metabolites stimulate calcium release from sea urchin egg microsomes desensitized to inositol trisphosphate. (англ.) // The Journal of biological chemistry. — 1987. — Vol. 262, no. 20. — P. 9561—9568. — PMID 3496336.
↑ Imai S., Armstrong C. M., Kaeberlein M., Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. (англ.) // Nature. — 2000. — Vol. 403, no. 6771. — P. 795—800. — DOI:10.1038/35001622. — PMID 10693811.
Литература |
- David E. Metzler. Biochemistry: The Chemical Reactions of Living Cells.. — 2nd edition. — Academic Press, 2003. — Т. 2. — 1973 с. — ISBN 978-0-1249-2541-0.
- David L. Nelson, Michael M. Cox. Lehninger Principles of biochemistry. — Fifth edition. — New York: W. H. Freeman and company, 2008. — 1158 p. — ISBN 978-0-7167-7108-1.
- Campbell N. A., Reece J. B., Urry L. A. e. a. Biology. 9th ed. — Benjamin Cummings, 2011. — 1263 p. — ISBN 978-0-321-55823-7.
- Кольман Я., Рём К.—Г. Наглядная биохимия. — 4-е изд.. — М.: БИНОМ. Лаборатория знаний, 2012. — 469 с. — ISBN 978-5-9963-0620-6.
- Биологическая химия с упражнениями и задачами / Под ред. С. Е. Северина. — М.: Издательская группа «ГЭОТАР-Медиа», 2011. — 624 с.
Коферменты | |
|---|---|
| Витамины |
|
| Не витамины |
|
| Минералы |
|
Эта статья входит в число хороших статей русскоязычного раздела Википедии. |


